Аквапорины вид транспорта: Презентация на тему: Виды транспорта
Атлас контактов
Информация о белок-белковых контактах
Субъединицы белка не связаны между собой ковалентно.
Однако в нем присутствуют немногочисленные солевые мостики (разновидность ионной связи между COO– и Nh4+ группами). Основной вклад в поддержание тетрамерной структуры вносят гидрофобные карманы и стекинг-взаимодействия.
Эти скрипты можно посмотреть в апплете ниже. Кстати, с помощью особых скриптов, мы посчитали, сколько есть разных вариантов стекинг-взаимодействий:
Phe-Phe – 27, His-Phe – 7, His-His – 0, Tyr-Phe – 8, Tyr-His – 0, Tyr-Tyr – 5, Trp-Phe – 12, Trp-His – 5, Trp-Tyr – 12, Trp-Trp – 3. Гидрофобные карманы искали при помощи Clud.
Водородные связи также вносят вклад в объединение субъединиц, но поскольку технические возможности JMol не позволяют автоматически их найти, мы показали как минимум одну вручную.
Координаты атомов на рисунке:
Белый [ILE]15:A.H/3 #57 48.115 37.977 41.335
Синий [ILE]15:A.N/3 #49 47.869 37.642 40. 583
583
Красный [GLY]11:A.O/4 #4 48.477 39.185 42.887
Плотность упаковки атомов в гидрофобном ядре
Мы взяли для рассмотрения ядро у Phe58, взятого в качестве MyResidue. В окружении этого аминокислотного остатка в основном присутствуют остатки лейцина, аланина и фенилаланина.
Атомы на расстоянии 1 Å от остатка MyResidue отсутствуют и присутствуют, начиная с 2 Å. C 7 Å атомы Phe58 уже практически полностью закрыты.
Для вычисления характерного расстояния между соседними не связанными ковалентно атомами в гидрофобном ядре было посчитано среднее значение длины среди 28 связей. Среднее расстояние равно 2,78 Å.
Для того, чтобы выяснить, сможет ли поместиться между соседними атомами ядра еще один, было высчитано среднее расстояние между границами Ван-дер-Ваальсовых радиусов.
Данная величина равна 0,63 Å, что говорит о том, что, например, молекула воды, которая имеет радиус в районе 1,4 Å, маловероятно сможет поместится между атомами гидрофобного ядра.
Previous Next
Информация о лиганд-белковых контаках
BOG
Отметим, что это молекулы растворителя, которыми удалось извлечь белок из мембраны, а не простетическая часть реального белка. Тем не менее, мы изучили, как он с ним связывается. Ионных, ковалентных взаимодействий и солевых мостиков ожидать не приходится. Но вот своим углеводородным хвостом он связывается с фенилаланином, то есть с помощью “гидрофобных” взаимодействий. Также BOG образует водородные связи с белком (см. скрипт).
Хлорид-ион
Функция хлорид-иона в аквапорине остается неясной. Более того, большая часть аквапоринов не пропускает их через себя, зато пропускает человеческий AQP6 [7].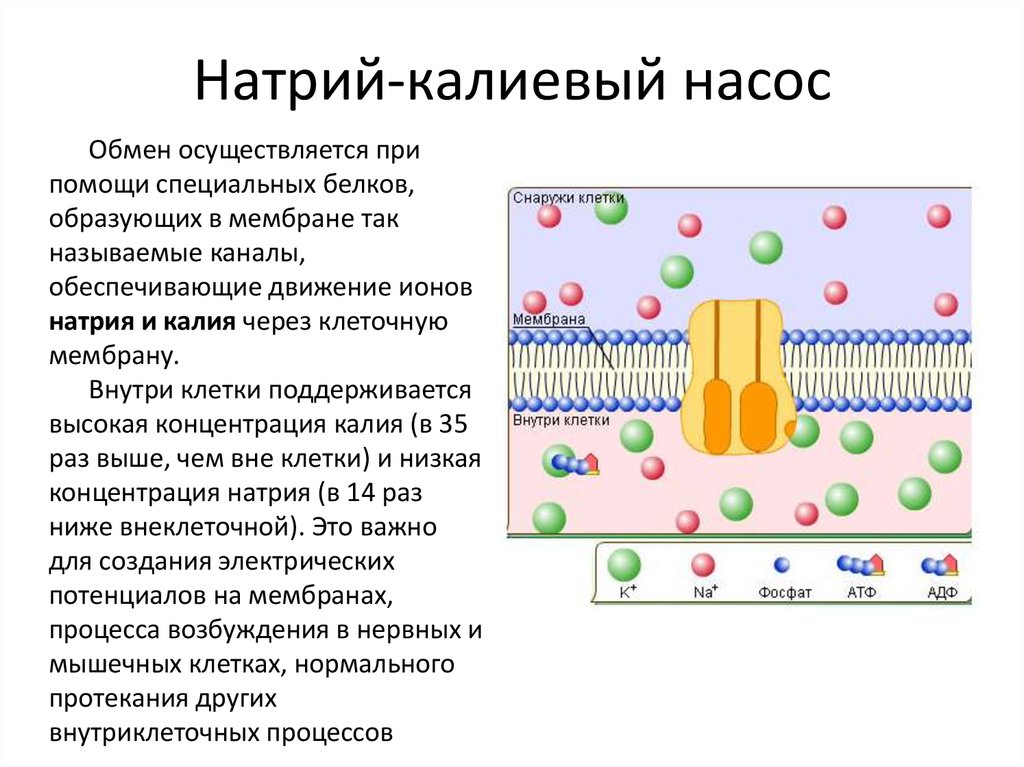 Один хлорид-ион находится прямо в канале и “закупоривает” отверстие.
Возможно, именно из-за того, что он там застрял, мембрана с аквапорином обладает свойством избирательной проницаемости.
Удерживается он в канале несколькими аргининами сверху, которые несут положительный заряд и несколькими триптофанами снизу.
Есть основания полагать, что аргинины там и нужны, чтобы задерживать отрицательно-заряженные ионы.
Остальные хлорид-ионы держатся засчет δ+ заряда NH группы пептидной связи. Все скрипты можно найти ниже в апплете.
Один хлорид-ион находится прямо в канале и “закупоривает” отверстие.
Возможно, именно из-за того, что он там застрял, мембрана с аквапорином обладает свойством избирательной проницаемости.
Удерживается он в канале несколькими аргининами сверху, которые несут положительный заряд и несколькими триптофанами снизу.
Есть основания полагать, что аргинины там и нужны, чтобы задерживать отрицательно-заряженные ионы.
Остальные хлорид-ионы держатся засчет δ+ заряда NH группы пептидной связи. Все скрипты можно найти ниже в апплете.
JMol апплет
Ссылки на пояснительные скрипты
1. Почему водородных связей между субъединицами нет
2. Почему нет ковалентных связей между субъединицами
3. Солевые мостики
4. Стекинг-взаимодействия
5. Какие есть стекинг-взаимодействия
6. Гидрофобные взаимодействия между мономерами
7. Гидрофобное ядро
8. Как связан BOG
9. Как связаны хлорид-ионы
Информация о положении в мембране
Чтобы предсказать по первичной последовательности, какие участки белка погружены в мембрану, а какие нет, мы воспользовались сервисом Transmembrane protein topology prediction with a hidden Markov model.
На оси Y отображены соответсвующие вероятности, а на оси Х – номер аминокислотного остатка белка. Красным цветом показаны предполагаемые трансмембранные части, розовым – части, обращеннные наружу, синим – части, обращенные внутрь.
Было предсказано 6 трансмембранных частей, 3 находящихся снаружи и 4 внутри, что соответствует действительности. Хотя можно заметить, что координаты участков отличаются в среднем на 4-5а.о., предсказание выполнено довольно точно.
Визуализация расположения белка в мембране
Для представления расположения аквапорина в мембране мы взяли существующую модель из банка PDB
Заключение
Для изучения был взят аквапорин Aqy1 Pichia Pastoris. В ходе исследования данного белка было выяснено, что он является тетрамером, т. е. состоит из 4 субъединиц-мономеров.
Также аквапорин имеет 2 низкомолекулярных лиганда (хлорид-ион и β-D-октил-глюкозид, которого находится по 3 в каждом мономере). Были изучены связи между ними.
Еще наша команда изучила гидрофобное ядро, и в ходе исследования были сделаны некоторые выводы по его плотности упаковки.
В ходе работы были также сделаны предсказания о положении белка в мембране, которые в дальнейшем оказались соответствующими действительности.
е. состоит из 4 субъединиц-мономеров.
Также аквапорин имеет 2 низкомолекулярных лиганда (хлорид-ион и β-D-октил-глюкозид, которого находится по 3 в каждом мономере). Были изучены связи между ними.
Еще наша команда изучила гидрофобное ядро, и в ходе исследования были сделаны некоторые выводы по его плотности упаковки.
В ходе работы были также сделаны предсказания о положении белка в мембране, которые в дальнейшем оказались соответствующими действительности.
Личный вклад
Владимиров Даниил исследовал белок-белковые взаимодействия, подготовил видеоролик и иллюстрации.
Петрухин Егор занимался изучением гидрофобного ядра, разработал соответствующие скрипты и иллюстрации.
Носкова Елизавета анализировала положение аквапорина в мембране, BOG- и белок-белковые взаимодействия, написала скрипты для этого.
Никитин Павел оформил отчёт и разместил его на своем сайте, а также изучал особенности хлорид-белковых взаимодействий.
Ссылки на источники
[1] Bonhivers M.
[2] Sidoux-Walter F., Pettersson N., Hohmann S. The Saccharomyces cerevisiae aquaporin Aqy1 is involved in sporulation //Proceedings of the National Academy of Sciences. – 2004. – Т. 101. – №. 50. – С. 17422-17427. doi: 10.1073/pnas.0404337101
[3] yeastgeonome.org (AQY1 / YPR192W Regulation)
[4] PDB in Europe
[5] Uniprot Aqy1 homologus
[6]. Дж. Эмсли. Элементы. 1993г
[7] Abir-Awan M. et al. Inhibitors of mammalian aquaporin water channels //International journal of molecular sciences. – 2019. – Т. 20. – №. 7. – С. 1589. doi: 10.3390/ijms20071589
Транспорт воды через аквапорины мембраны клетки
3 функции- 3.1 Функции у животных
- 3.2 Функции в растениях
- 3.3 Функции в микроорганизмах
С базовыми знаниями в области физиологии и механизмов, которые перемещаются растворенные через мембрану (активной и пассивной), мы могли бы чувствовать, что водный транспорт не является проблемой, и из клетки путем простой диффузии.
Эта идея управлялась в течение многих лет. Тем не менее, некоторые исследователи мелькают существование любого канала водного транспорта, так как в некоторых типах клеток с высокой проницаемостью для воды (например, почек, например), диффузия не достаточен механизма для объяснения транспорта вода.
Врач и исследователь Питер Агре открыл эти белковые каналы в 1992 году, работая с мембраной эритроцитов. Благодаря этому открытию он получил (вместе со своими коллегами) Нобелевскую премию в 2003 году. Этот первый аквапорин получил название «Аквапорин 1»..
структураФорма аквапорина напоминает песочные часы с двумя симметричными половинами, ориентированными в противоположных направлениях. Эта структура пересекает двойную липидную мембрану клетки.
Следует отметить, что форма аквапоринов очень специфична и не похожа ни на какие другие типы белков, которые пересекают мембрану..
Аминокислотные последовательности являются преимущественно полярными. Трансмембранные белки характеризуются наличием сегмента, богатого альфа-спиральными сегментами. Однако у аквапоринов нет таких регионов.
Трансмембранные белки характеризуются наличием сегмента, богатого альфа-спиральными сегментами. Однако у аквапоринов нет таких регионов.
Благодаря использованию современных технологий, стало возможным уточнить в деталях структуру порина: мономеры от 24 до 30 кДа, состоящий из шести геликоидов segmentes двух небольших сегментов, окружающих цитоплазму и соединены с малыми порами.
Эти мономеры собраны в группу из четырех единиц, хотя каждый может работать независимо. У маленьких пропеллеров есть некоторые законсервированные мотивы, включая NPA.
У некоторых аквапоринов, обнаруженных у млекопитающих (AQP4), наблюдаются более высокие скопления, которые образуют надмолекулярные кристаллические структуры..
Для транспортировки воды внутренняя часть белка является полярной, а внешняя – неполярной, в отличие от обычных глобулярных белков.
Функция аквапоринов заключается в том, чтобы опосредовать перенос воды внутрь клетки в ответ на осмотический градиент.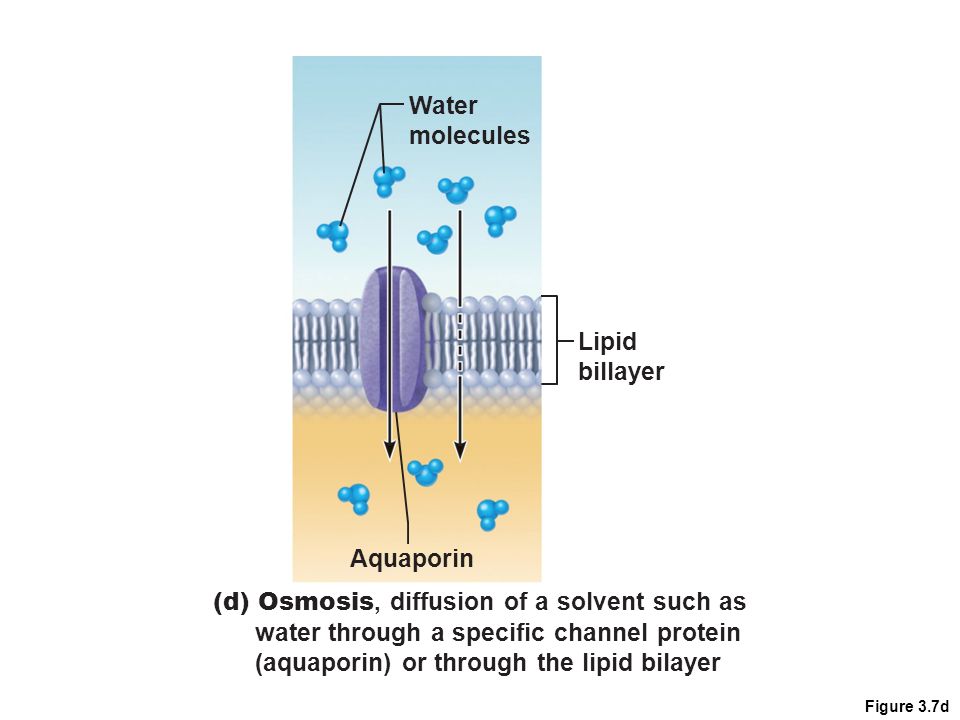 Для этого не требуется никакой дополнительной силы или перекачки: вода поступает в клетку и покидает ее посредством осмоса, опосредованного аквапорином. Некоторые варианты также несут молекулы глицерина.
Для этого не требуется никакой дополнительной силы или перекачки: вода поступает в клетку и покидает ее посредством осмоса, опосредованного аквапорином. Некоторые варианты также несут молекулы глицерина.
Чтобы осуществить этот транспорт и существенно увеличить водопроницаемость, клеточная мембрана изобилует молекулами аквапорина, порядка плотности 10000 квадратных микрометров..
Функции у животных
Перевозка воды жизненно важна для организмов. Давайте возьмем пунктуальный пример почек: они должны ежедневно фильтровать огромное количество воды. Если этот процесс не происходит должным образом, последствия будут фатальными.
Кроме того, концентрация мочи, аквапорины участвуют в общем гомеостазе организма жидкости в функции мозга, секреции желез, увлажнения кожи, мужской фертильности, зрение, слух это лишь некоторые процессы биологический.
В экспериментах, проведенных на мышах, был сделан вывод о том, что они также участвуют в миграции клеток, роль, которая далека от водного транспорта. .
.
Функции в растениях
Аквапорины в основном разнообразны в царстве растений. В этих организмах опосредуют важные процессы, такие как пот, размножение, обмен веществ.
Кроме того, они играют важную роль в качестве адаптивного механизма в средах, где условия окружающей среды не являются оптимальными.
Функции в микроорганизмах
Хотя аквапорины присутствуют в микроорганизмах, конкретная функция еще не найдена.
Главным образом по двум причинам: высокое отношение объема поверхности микробов предполагает быстрое осмотическое равновесие (исключая необходимость в аквапоринах), а исследования делеций микробов не дали четкого фенотипа..
Тем не менее, предполагается, что аквапорины могут обеспечить некоторую защиту от последовательных событий замерзания и оттаивания, поддерживая водопроницаемость в мембранах при низких температурах..
типМолекулы аквапорина известны в различных линиях, как у растений, так и у животных и у менее сложных организмов, и они очень похожи друг на друга – тогда мы предполагаем, что они появились на ранних стадиях эволюции.![]() .
.
Растения найдены в 50 различных молекулах, в то время как млекопитающие обладают только 13, распределенными по различным тканям, включая эндотелиальную эпителиальную ткань и почки, легкие, экзокринные железы и органы пищеварения, связанную с.
Тем не менее, аквапорины также выражены в тканях, которые не имеют очевидную и прямую связь с транспортировкой жидкостей в организме, как и в астроцитах центральной нервной системы и в некоторых регионах глаз, такие, как роговицы и ресничный эпителий.
Есть аквапорины в грибковой мембране бактерий (как Кишечная палочка) и в мембранах органелл, таких как хлоропласты и митохондрии.
Медицинские патологии, связанные с аквапоринамиУ пациентов, у которых есть некоторый дефект в последовательности аквапорина 2, присутствующего в клетках почки, они должны потреблять более 20 литров воды, чтобы поддерживать себя в увлажненном состоянии. В этих медицинских случаях адекватная концентрация мочи отсутствует.
Противоположный случай также приводит к интересному клиническому случаю: выработка избытка аквапорина 2 приводит к чрезмерной задержке жидкости у пациента.
В периоды беременности происходит увеличение синтеза аквапоринов. Этот факт объясняет задержку жидкости, которая часто встречается у будущих мам. Точно так же отсутствие аквапорина 2 было связано с появлением определенного типа диабета..
Транспорт воды через аквапорины мембраны клетки
СПРАВОЧНИК НЕВРОЛОГА
ВВЕДЕНИЕ
Вода составляет примерно 70% массы большинства живых организмов. Однако содержание ее внутри и вне клетки различно, и, как ни странно, клетка вынуждена строго регулировать поступление этого «универсального» растворителя. Этой цели служат специальные белки клеточной мембраны – аквапорины.
Зачем же клетке так тщательно заботиться о количестве такого, казалось бы, безвредного вещества, как вода? Оказывается, регуляция водного баланса внутри клеток влияет на их способность выполнять специфическую функцию, необходимую для жизни всего организма. Регулируя поступление воды и тем самым свой объем, нервные клетки, например, влияют на состояние ионных каналов, передающих нервные импульсы .
Регулируя поступление воды и тем самым свой объем, нервные клетки, например, влияют на состояние ионных каналов, передающих нервные импульсы .
Концентрация солей, жиров, белков и углеводов внутри клетки превышает таковую снаружи, а это значит, что относительное количество воды в ней меньше, чем в тканевых жидкостях. Клетка испытывает постоянное давление воды, которому должна сопротивляться в течение всей жизни. Если сравнить под микроскопом мертвую и живую клетки из одной и той же ткани, можно сразу же заметить, что мертвая значительно больше: это вода вошла внутрь и «раздула» ее ( Laesus De Liro – аналогия – отек ткани, клеток, головного мозга на фоне острой очаговой ишемии [инфаркта ткани мозга] или диффузный отек [набухание] головного мозга на фоне острых метаболических нарушений и т.д.).
Хотя некоторое количество воды способно пассивно (простой диффузией) проникать через липидные слои, составляющие основу клеточных мембран (а также путем котранспорта через транспортеры электролитов и глюкозы), скорость и интенсивность такого водообмена явно недостаточна. Следовательно, клетке необходим специальный механизм для регуляции водных потоков. Такой механизм должен к тому же быть очень специфичным, чтобы вместе с молекулой воды не проникали вредные водорастворимые (гидрофильные) соединения, которые могут повредить ДНК, клеточные белки, либо блокировать внутриклеточные реакции.
Следовательно, клетке необходим специальный механизм для регуляции водных потоков. Такой механизм должен к тому же быть очень специфичным, чтобы вместе с молекулой воды не проникали вредные водорастворимые (гидрофильные) соединения, которые могут повредить ДНК, клеточные белки, либо блокировать внутриклеточные реакции.
Удовлетворению этих потребностей служит семейство мембранных каналообразующих белков (специфические системы транспорта воды), которые обеспечивают быстрый транспорт воды через биологические мембраны. Эти белки называют аквапоринами, они обнаружены у всех живых организмов. Открыты аквапорины были 30 лет назад, и с тех пор исследователи со всего мира пристально изучают их структуру и функции.
СТРУКТУРА АКВАПОРИНОВ
Существование водных каналов, пронизывающих мембраны клеток, постулировалось различными авторами с 1950-х годов, но молекулярные механизмы трансмембранного движения воды оставались неясными до работ P. Agre, который в 1988 г. открыл внутримембранный белок массой 28 kDa, идентифицированный как аквапорин 1 (AQP1). В 2003 г. P. Agre за исследование водных каналов была присуждена Нобелевская премия. Сегодня известно около 200 различных аквапоринов (Agre P. et al., 2003), формирующих водные каналы в клеточных мембранах всех живых существ. Нумерация аквапоринов, начиная с AQP 1, связана с последовательностью их открытия.
В 2003 г. P. Agre за исследование водных каналов была присуждена Нобелевская премия. Сегодня известно около 200 различных аквапоринов (Agre P. et al., 2003), формирующих водные каналы в клеточных мембранах всех живых существ. Нумерация аквапоринов, начиная с AQP 1, связана с последовательностью их открытия.
Аквапорины – гидрофобные трансмембранные белки (отдельные мономеры) массой от 26 до 34 kDa, формирующие гомотетрамеры. Отдельные мономеры состоят в среднем из 270 аминокислотных остатков (Титовец, 2007). Каждый мономер формирует отдельную водную пору. Проницаемость мономеров не меняется при их ассоциации в [гомо]тетрамер. В центре тетрамера также имеется пора, роль которой окончательно не установлена. Имеются предположения, что пора представляет собой лиганд-зависимый ионный канал.
Полипептидная цепь мономера пронизывает мембрану шесть раз, N- и C-концы обращены внутрь клетки. Концевые участки являются мишенями посттрансляционной модификации. Участки цепи, находящиеся в мембране (т.![]() е. пронизывающие мембрану шесть раз) образуют 6 α-спиралей, которые наклонены под углом примерно 25 градусов по отношению к плоскости мембраны. Между N- и C-половинами молекулы наблюдается высокая степень гомологии, что может быть следствием древней дупликации аквапоринового гена. На соединительных петлях B и E практически у всех представителей семейства аквапоринов имеется консервативный мотив, состоящий из 3 аминокислот: аспарагина, пролина и аланина (NPA-мотив). Петли B и Е несут короткие α-спиральные домены, которые погружены в мембрану. Эти домены принимают участие в формировании высокоселективного канала для воды (проницаемость аквапоринов высока: 3х10*9 молекул воды в секунду на мономер, но при этом очень избирательна). Данная модель мономера аквапоринов получила название модели «песочных часов» (hourglass model).
е. пронизывающие мембрану шесть раз) образуют 6 α-спиралей, которые наклонены под углом примерно 25 градусов по отношению к плоскости мембраны. Между N- и C-половинами молекулы наблюдается высокая степень гомологии, что может быть следствием древней дупликации аквапоринового гена. На соединительных петлях B и E практически у всех представителей семейства аквапоринов имеется консервативный мотив, состоящий из 3 аминокислот: аспарагина, пролина и аланина (NPA-мотив). Петли B и Е несут короткие α-спиральные домены, которые погружены в мембрану. Эти домены принимают участие в формировании высокоселективного канала для воды (проницаемость аквапоринов высока: 3х10*9 молекул воды в секунду на мономер, но при этом очень избирательна). Данная модель мономера аквапоринов получила название модели «песочных часов» (hourglass model).
Подробнее о строении и функционировании аквапоринов в следующих источниках:
статья «Лауреаты Нобелевской премии 2003 года по химии (Нобелевская премия по химии за 2003 г.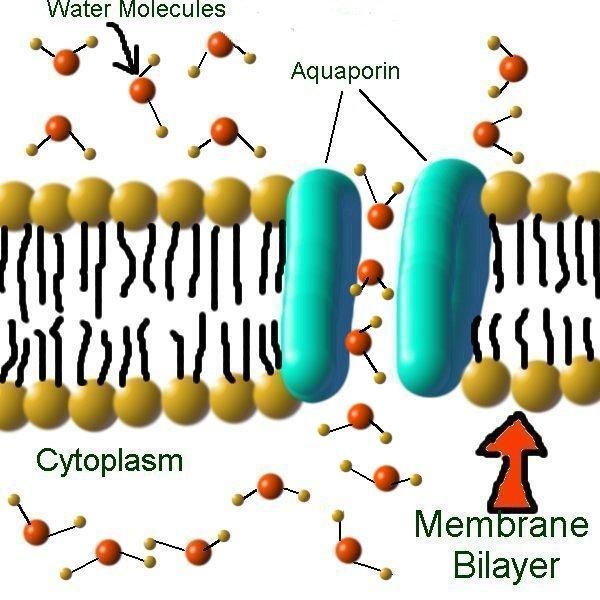 присуждена Р. Мак-Киннону и П. Эгру за открытия, касающиеся переноса ионов и молекул воды через клеточную мембрану)» Белянова Л.П., кандидат химических наук, Москва (журнал «Природа» №1, 2004) [читать] или [читать];
присуждена Р. Мак-Киннону и П. Эгру за открытия, касающиеся переноса ионов и молекул воды через клеточную мембрану)» Белянова Л.П., кандидат химических наук, Москва (журнал «Природа» №1, 2004) [читать] или [читать];
статья (обзор) «Аквопорины и гипервазопрессинеия» И.Н. Кабанов, Л.П. Чурилов; Кафедра патологии Медицинского факультета Санкт-Петербургского государственного университета (журнал «Медицина XXI век» № 5(6), 2007) [читать]
В отношении функциональной активности аквапоринов можно сделать следующие выводы (Шапигузов, 2004): [ 1 ] В присутствии аквапоринов скорость транспорта воды и некоторых других неэлектролитов через мембрану возрастает, а энергия активации этого процесса снижается. [ 2 ] Транспорт через аквапорины протекает с одинаковой скоростью в обоих направлениях при одинаковой разности химических потенциалов переносимого вещества по разные стороны мембраны. [ 3 ] Транспорт через аквапорины является пассивным. [ 4 ] Для аквапоринов характерна селективность. Транспорт незаряженной молекулы через пору мономера не сопровождается переносом ионов, в том числе и протонов. [ 5 ] Некоторые аквапорины ингибируются соединениями тяжелых металлов (Hg, Ag, Au, Cu, Ni).
Классификация аквапоринов основана на их специфичности по отношению к транспортируемым молекулам и аминокислотной последовательности. Первая группа включает собственно аквапорины специфичные по отношению к воде. К этой группе относятся следующие аквапорины млекопитающих AQP0, AQP1, AQP2, AQP4, AQP5, AQP6 и AQP8. Вторая группа включает акваглицеропорины. Различные представители этой группы в разной степени проницаемы для воды, но дополнительно проницаемы для других малых неэлектролитов, в первую очередь для глицерола и мочевины. К этой группе относятся 4 аквапорина млекопитающих (AQP3, AQP7, AQP9 и AQP10) (Титовец, 2007). Транспортные свойства AQP11 и AQP12 (супераквапоринов) к настоящему времени исследованы недостаточно. В головном мозге определяются AQP1,AQP3, AQP4, AQP9, AQP11.
подробнее в статье «Роль аквапоринов в транспорте воды через биологические мембраны (обзор литературы)» А.В. Крысова, В.И. Циркин, А.А. Куншин; Вятский государственный гуманитарный университет; Казанский государственный медицинский университет (журнал «Вятский медицинский вестник» №2, 2012) [читать]
Нарушения экспрессии аквапоринов важны в этиологии и патогенезе большого числа патологических процессов. Дефект различных аквапоринов является причиной (этиологическим фактором) нарушения водного баланса мозга , некоторых случаев бронхиальной астмы, нарушения реабсорбции воды в кишечнике и почках, глаукомы, катаракты, нарушения продукции слюны, недостаточного потоотделения и так далее.
АКВАПОРИНЫ И ЦЕРЕБРАЛЬНЫЙ ОТЕК
За последнее десятилетие в области изучения патогенеза церебрального отека был достигнут значительный прогресс благодаря открытию аквапоринов и выяснению их роли в водном обмене структур головного мозга в норме и при патологии. Ранее при описании механизмов отека головного мозга речь шла, в общем, о накоплении воды вследствие нарушения проницаемости гематоэнцефалического барьера, деструкции клеточных мембран и т.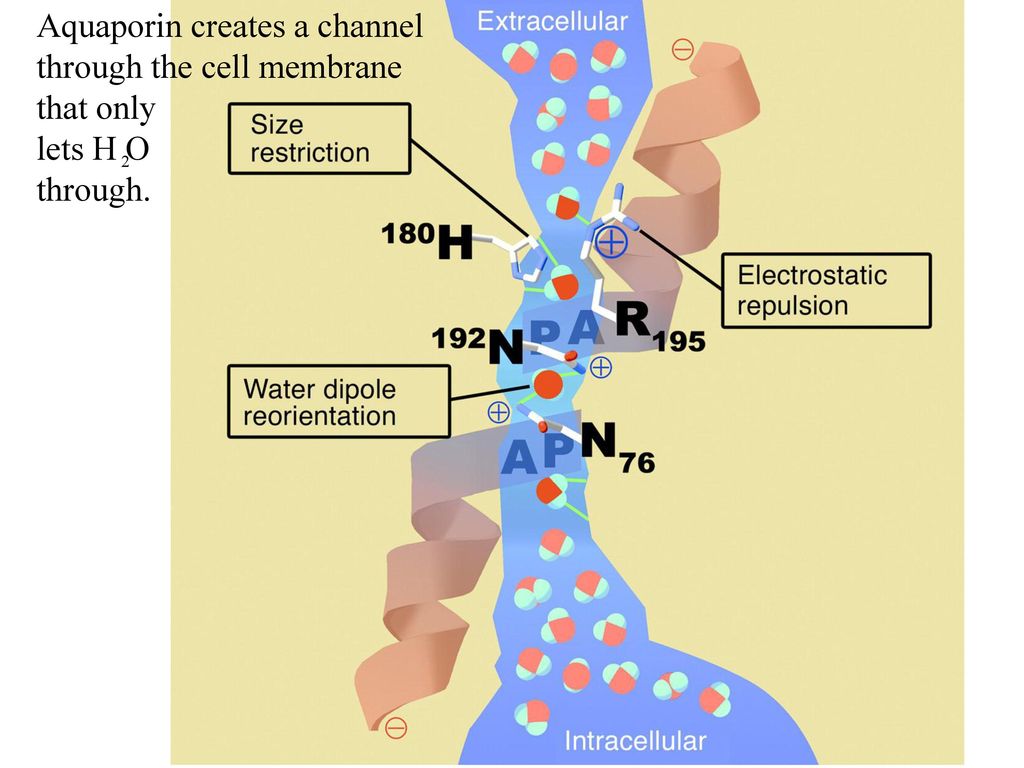 д., при этом не было представления о конкретных молекулярных механизмах нарушения водного обмена. Согласно новому фундаментальному знанию, водный обмен головного мозга осуществляется при непосредственном участии аквапоринов AQP1, AQP4 и AQP9. Эти водные каналы поддерживают водный гомеостаз, пространственно организуют и обеспечивают облегченный транспорт воды между основными водными компартментами головного мозга: [ 1 ] водой крови и жидкостью паренхимы головного мозга; [ 2 ] внутриклеточной и внеклеточной жидкостью; [ 3 ] цереброспинальной жидкостью (ЦСЖ) желудочков и водой субарахноидального пространства. Основная нагрузка при этом ложится на аквапорины AQP1 и AQP4, однако их функциональная роль в физиологических и патологических условиях различна. Все аквапорины свободно переносят воду как в прямом, так и в обратном направлении в зависимости от направления вектора осмотического или гидростатического давления. От уровня их экспрессии в плазматической мембране клетки, а также от уровня их активности зависит величина водного потока. В отличие от AQP4 аквапорины AQP1 и AQP9 являются переносчиками не только воды, но и газов, а также других важных биорегуляторов и метаболитов.
д., при этом не было представления о конкретных молекулярных механизмах нарушения водного обмена. Согласно новому фундаментальному знанию, водный обмен головного мозга осуществляется при непосредственном участии аквапоринов AQP1, AQP4 и AQP9. Эти водные каналы поддерживают водный гомеостаз, пространственно организуют и обеспечивают облегченный транспорт воды между основными водными компартментами головного мозга: [ 1 ] водой крови и жидкостью паренхимы головного мозга; [ 2 ] внутриклеточной и внеклеточной жидкостью; [ 3 ] цереброспинальной жидкостью (ЦСЖ) желудочков и водой субарахноидального пространства. Основная нагрузка при этом ложится на аквапорины AQP1 и AQP4, однако их функциональная роль в физиологических и патологических условиях различна. Все аквапорины свободно переносят воду как в прямом, так и в обратном направлении в зависимости от направления вектора осмотического или гидростатического давления. От уровня их экспрессии в плазматической мембране клетки, а также от уровня их активности зависит величина водного потока. В отличие от AQP4 аквапорины AQP1 и AQP9 являются переносчиками не только воды, но и газов, а также других важных биорегуляторов и метаболитов.
AQP1 представляет собой количественно преобладающий водный канал эпителия хориоидального сплетения. Здесь он экспрессируется в апикальной мембране клеток хориоидального сплетения и играет главную роль в продуцировании ЦСЖ. Считается, что в норме в остальной части головного мозга AQP1 отсутствует или его экспрессия невысока. Вместе с тем биохимические и гистохимические исследования по идентификации AQP1 на секционном материале нормального мозга человека позволили обнаружить относительно небольшое количество AQP1 примерно в 33% микрососудов головного мозга. Интересно отметить, что в эндотелии кровеносных сосудов вне головного мозга AQP1 экспрессируется в большом количестве и определяет интенсивность гистогематического обмена воды. В целом вопрос об уровне его экспрессии в клетках эндотелия микрососудов головного мозга остается открытым. По-видимому, наличие неоднозначных данных о распределении AQP1 в тканях головного мозга связано с тем, что многие патологические состояния, начиная с гипоксии, сопровождаются активацией экспрессии этого канала в эндотелии микрососудов. Так, аквапорин AQP1 экспрессируется в большом количестве в эндотелии микрососудов астроцитомы человека и митостатических карциномах. Он обнаруживается также в эндотелии микрососудов глиобластом. Экспрессия AQP1 возрастает при опухолях хориоидального сплетения, что сопровождается повышением продуцирования ЦСЖ.
По-видимому, наличие неоднозначных данных о распределении AQP1 в тканях головного мозга связано с тем, что многие патологические состояния, начиная с гипоксии, сопровождаются активацией экспрессии этого канала в эндотелии микрососудов. Так, аквапорин AQP1 экспрессируется в большом количестве в эндотелии микрососудов астроцитомы человека и митостатических карциномах. Он обнаруживается также в эндотелии микрососудов глиобластом. Экспрессия AQP1 возрастает при опухолях хориоидального сплетения, что сопровождается повышением продуцирования ЦСЖ.
AQP4 является самым активным водным каналом. В центральной нервной системе он обнаруживается в астроцитах всех отделов головного и спинного мозга и клетках эпендимы, выстилающей желудочки. Его экспрессия избирательна, вследствие чего AQP4 локализуется лишь в определенных субклеточных структурах. В астроцитах, где отмечается высокополяризованное субклеточное распределение AQP4, он сосредоточен в оконечных ножках астроцитов и базолатеральных областях плазматической мембраны ограничивающей глии, т. е. на поверхностях разделов кровь-паренхима или ЦСЖ-паренхима головного мозга. Обычно этот аквапорин организован в так называемые ортогональные структуры. Концевые отростки ножек астроцитов, где сосредоточен AQP4, находятся в тесном контакте с мозговыми микрососудам, а также с синапсами. Именно AQP4 в концевых отростках астроглиальных клеток определяет проницаемость к воде гематоэнцефалического барьера.
е. на поверхностях разделов кровь-паренхима или ЦСЖ-паренхима головного мозга. Обычно этот аквапорин организован в так называемые ортогональные структуры. Концевые отростки ножек астроцитов, где сосредоточен AQP4, находятся в тесном контакте с мозговыми микрососудам, а также с синапсами. Именно AQP4 в концевых отростках астроглиальных клеток определяет проницаемость к воде гематоэнцефалического барьера.
AQP4 обеспечивает быстрое локальное перемещение воды из периваскулярного пространства (вследствие увеличения уровня экспрессии AQP4), что лежит в основе механизма контроля как собственно объема периваскулярного пространства, так и регуляции скорости церебральной перфузии (например, при астроцитоме уровни экспрессии AQP4 в перитуморозной области, по данным магнитнорезонансной томографии [МРТ], коррелируют с зоной распространения отека). Наличие коррелятивных взаимоотношений между экспрессией AQP4 и выраженностью отека установлено также при ишемии, травме головного мозга и субарахноидальных кровоизлияниях. Важная роль AQP4 в патогенезе церебрального отека была экспериментально обоснована и однозначно доказана многочисленными исследованиями. При этом AQP4 выполняет бимодальную функцию в патогенезе церебрального отека, и его конкретная роль зависит от того, какой отек преобладает – вазогенный или цитотоксический. При вазогенном отеке процесс реабсорбции жидкости может быть ускорен путем повышения уровня экспрессии AQP4, а развитие цитотоксического отека на ранних стадиях ишемии может быть предотвращено путем понижения активности этого аквапорина.
Обратите внимание! Следует помнить о том, что высокоспецифические аутоантитела к AQP4 (NMO-IgG) вызывают демиелинизирующие поражения ЦНС, при которых клинически наблюдается преимущественное поражение зрительных нервов и/или спинного мозга («спектр оптикомиелит-ассоциированных расстройств» – neuromyelitis optica spectrum disorders, или NMOSD). Также AQP4 учавствует в функционировании глимфатической система головного мозга.
статью «Глимфатическая система головного мозга: функциональная анатомия и клинические перспек-тивы» Николенко В. Н., Оганесян М.В., Яхно Н.Н., Орлов Е.А., Порубаева Э.Э., Попова Е.Ю.; ФГАОУ ВО «Первый Московский государственный медицинский университет имени И.М. Сеченова» МЗ РФ, Москва; ФГБОУ ВО «Московский государственный университет имени М.В. Ломоносова», Москва (журнал «Неврология, нейропсихиатрия, психосоматика» №4, 2018) [читать]
Н., Оганесян М.В., Яхно Н.Н., Орлов Е.А., Порубаева Э.Э., Попова Е.Ю.; ФГАОУ ВО «Первый Московский государственный медицинский университет имени И.М. Сеченова» МЗ РФ, Москва; ФГБОУ ВО «Московский государственный университет имени М.В. Ломоносова», Москва (журнал «Неврология, нейропсихиатрия, психосоматика» №4, 2018) [читать]
Роль AQP9, который экспрессируется в астроцитах и катехоламинэргических нейронах, не ограничивается только функцией транспорта воды. Этот аквапорин принимает участие в энергетическом обмене головного мозга, обеспечивая перенос лактата, глицерина и других энергетических субстратов. Следует отметить, что экспрессия AQP9 зависит от концентрации инсулина. При ишемии уровень экспрессии этого аквапорина повышается, и имеются указания на то, что AQP9 может принимать участие в утилизации лактата, накапливающегося при ишемии головного мозга. В отличие от AQP4, AQP9 не отводится основная роль в патогенезе отека.
Подробнее об аквапоринах и церебральном отеке в следующих источниках:
статья «Церебральный отек и современные направления его лечения» Э. П. Титовец, А.Ф. Смеянович; Республиканский научно-практический центр неврологии и нейрохирургии, Минск, Беларусь (журнал «Вестник национальной академии наук Белоруссии» №1, 2011) [читать];
П. Титовец, А.Ф. Смеянович; Республиканский научно-практический центр неврологии и нейрохирургии, Минск, Беларусь (журнал «Вестник национальной академии наук Белоруссии» №1, 2011) [читать];
статья «Клиническая патофизиология отека головного мозга (часть 1)» А.А. Задворнов, А.В. Голомидов, Е.В. Григорьев; ГАУЗ КО «Областная детская клиническая больница», г. Кемерово; ФГБУ «НИИ комплексных проблем сердечно-сосудистых заболеваний», г. Кемерово (журнал «Вестник анестезиологии и реаниматологии» №3, 2017) [читать];
статья «Клиническая патофизиология отека головного мозга (часть 2)» А.А. Задворнов, А.В. Голомидов, Е.В. Григорьев; ГАУЗ КО «Областная детская клиническая больница», г. Кемерово; ФГБУ «НИИ комплексных проблем сердечно-сосудистых заболеваний», г. Кемерово (журнал «Вестник анестезиологии и реаниматологии» №4, 2017) [читать];
[для подписчиков журнала «Вестник национальной академии наук Белоруссии»] статья «Коррекция нарушений церебрального водного обмена» Э. П. Титовец, А.Ф. Смеянович, П.В. Козич; Республиканский научно-практический центр неврологии и нейрохирургии, Минск, Беларусь (журнал «Вестник национальной академии наук Белоруссии» №1, 2019) [перейти]
П. Титовец, А.Ф. Смеянович, П.В. Козич; Республиканский научно-практический центр неврологии и нейрохирургии, Минск, Беларусь (журнал «Вестник национальной академии наук Белоруссии» №1, 2019) [перейти]
Механизм транспорта воды через мембраны.
В настоящее время растительную клетку рассматривают не только как осмотическую систему. По этому поступление и выход воды из клетки зависит не только от градиента водного потенциала – соотношения активности воды (или водного потенциала) внутри клетки и в растворе, но и от проницаемости плазмалеммы и тонопласта для входа воды.
Молекулы воды могут преодолевать биологические мембраны двумя путями: 1) через липидный бислой; 2) через поры, образуемыми специфическими белками – акавапоринами. Хотя молекула воды является диполем, она электронетральна и вследствие этого может в отличие от ионов диффундировать через липидную фазу мембраны. Долгое время путь воды через липидный бислой рассматривался как единственный.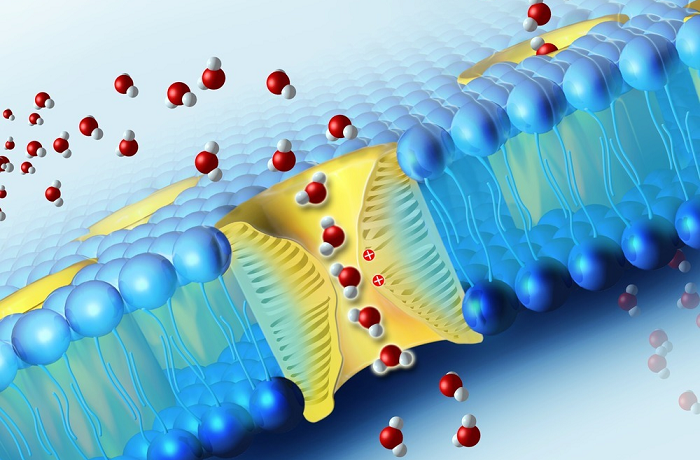 Однако известны физиологические процессы, связанные с интенсивной «переброской» больших объемов воды или быстрым изменением водной проницаемости мембран. Эти явления невозможно объяснить диффузией воды через липиды, что заставило предположить существование переносчиков воды белковой природы. Наличие в мембранах пор, пропускающих молекулы воды, стали предполагать, начиная с 1960-х годов. Позднее существование таких белковых пор было доказано, и они получили название водных каналов – аквапоринов.
Однако известны физиологические процессы, связанные с интенсивной «переброской» больших объемов воды или быстрым изменением водной проницаемости мембран. Эти явления невозможно объяснить диффузией воды через липиды, что заставило предположить существование переносчиков воды белковой природы. Наличие в мембранах пор, пропускающих молекулы воды, стали предполагать, начиная с 1960-х годов. Позднее существование таких белковых пор было доказано, и они получили название водных каналов – аквапоринов.
Студопедия рекомендует:
Принципы уголовного процесса Стоит прочитать соответствующую главу учебника Принципы уголовного процесса &ndash.
Трудовой потенциал: понятие, структура и показатели Для определения возможности участия человека в экономических и управленческих процессах на предприятии обычно используют такие.
Анализ производства и реализации продукции В настоящее время предприятия самостоятельно планируют свою деятельность и определяют перспективы развития исходя из спроса на.
Понятие о группе в психологии, классификация групп Группа – ограниченная в размерах общность людей, выделяемая из социального целого на основе определённых признаков.
Выпишите рецепты по образцу 1.Возьми: горькой настойки настойки ревеня по 15 г Смешай. Выдай. Обозначь: Rp.: Tincturae amarae Tincturae Rhei ana 15.
Жизнь – это не просто годы, посвященные ублажению самих себя и собственной карьере. Это привилегия, ответственность, необходимость жить, следуя за высшим призванием. © Элизабет Доул ==> читать все изречения.
Аквапорины – естественные проводники влаги и защитники кожи
Читайте также:
- Синдром Унны (Unna)
- КТ при гемангиоме толстой кишки
- Травма ногтей на пальцах рук и ног
- Вторичные опухоли орбиты: причины, диагностика, лечение
- Транспозиция главных сосудов плода. Диагностика транспозиции главных сосудов
Аквапорины
VIVO Патофизиология
Другие темы
Вода проникает через клеточные мембраны двумя путями: путем диффузии через липидный бислой и через водные каналы, называемые аквапоринами. Функциональная характеристика первого аквапорина была сообщена в 1992 г., но предполагалось, что водные каналы существовали задолго до этого времени, поскольку осмотическая проницаемость некоторых типов эпителиальных клеток была слишком велика, чтобы ее можно было объяснить простой диффузией через плазматическую мембрану.
Функциональная характеристика первого аквапорина была сообщена в 1992 г., но предполагалось, что водные каналы существовали задолго до этого времени, поскольку осмотическая проницаемость некоторых типов эпителиальных клеток была слишком велика, чтобы ее можно было объяснить простой диффузией через плазматическую мембрану.
Один человеческий канал аквапорина-1 способствует транспортировке воды со скоростью примерно 3 миллиарда молекул воды в секунду. Такой транспорт оказывается двунаправленным в соответствии с преобладающим осмотическим градиентом.
Классические аквапорины переносят свободную от растворенных веществ воду через клеточные мембраны; они представляют собой эксклюзивные водные каналы и не проникают через мембраны до ионов или других малых молекул. Некоторые аквапорины, известные как акваглицеропорины , транспортируют воду, глицерин и несколько других небольших молекул.
Семейство аквапоринов
На сегодняшний день идентифицировано более 10 различных аквапоринов млекопитающих, и предполагается, что существуют дополнительные представители. Близкородственные белки водных каналов были выделены из растений, насекомых и бактерий. Аквапорин-1 из эритроцитов человека был обнаружен первым и, вероятно, лучше всего изучен.
Близкородственные белки водных каналов были выделены из растений, насекомых и бактерий. Аквапорин-1 из эритроцитов человека был обнаружен первым и, вероятно, лучше всего изучен.
Графики гидрофобности их аминокислотных последовательностей показывают, что аквапорины имеют шесть трансмембранных сегментов, как показано на модели аквапорина-1 справа.
На основании исследований аквапорина-1 выяснилось, что аквапорины существуют в плазматической мембране в виде гомотетрамеров. Каждый мономер аквапорина содержит две полупоры, которые складываются вместе, образуя канал для воды.
Различные аквапорины имеют разные модели гликозилирования. В случае аквапорина-1 пептидный скелет составляет примерно 28 кДа, а масса гликозилированных форм варьируется от 40 до 60 кДа. Большинство аквапоринов имеют мотив фосфорилирования протеинкиназы А в одной из цитоплазматических петель, и предполагается, что дифференциальное фосфорилирование придает молекуле регуляторную функцию.
Паттерны экспрессии аквапоринов
Каждый из аквапоринов имеет уникальный характер экспрессии в тканях и во время развития. Краткое изложение этих атрибутов и некоторых важных потенциальных или известных функций представлено в следующей таблице:
Краткое изложение этих атрибутов и некоторых важных потенциальных или известных функций представлено в следующей таблице:
| | Основные сайты экспрессии | Комментарии |
|---|---|---|
| Аквапорин-0 | Глаз: клетки волокон хрусталика | Баланс жидкости в линзе |
| Аквапорин-1 | Эритроциты | Осмотическая защита |
| Почка: проксимальный каналец | Концентрация мочи | Глаз: реснитчатый эпителий | Производство водянистой влаги | Головной мозг: сосудистое сплетение | Производство спинномозговой жидкости | Легкие: альвеолярные эпителиальные клетки | Состояние гидратации альвеол |
| Аквапорин-2 | Почки: собирательные трубочки | Опосредует активность антидиуретического гормона |
| Аквапорин-3 * | Почки: собирательные трубочки | Реабсорбция воды в кровь |
| Трахея: эпителиальные клетки | Секреция воды в трахею | |
| Аквапорин-4 | Почки: собирательные трубочки | Реабсорбция воды |
| Головной мозг: эпендимальные клетки | Баланс жидкости для спинномозговой жидкости | |
| Головной мозг: гипоталамус | Функция осмосенсинга? | |
| Легкое: бронхиальный эпителий | Бронхиальная секреция | |
| Аквапорин-5 | Слюнные железы | Производство слюны |
| Слезные железы | Производство слез | |
| Аквапорин-6 | Почки | Очень низкая водопроницаемость; функция? |
| Аквапорин-7 * | Жировые клетки | Транспортирует глицерин из адипоцитов |
| Яички и сперматозоиды | | |
| Аквапорин-8 | Семенники, поджелудочная железа, печень, другие | |
| Аквапорин-9 * | Лейкоциты | |
| * акваглицеропорин | ||
Должно быть ясно, что аквапорины очень широко распространены, а также что разные аквапорины имеют разные функционально важные особенности.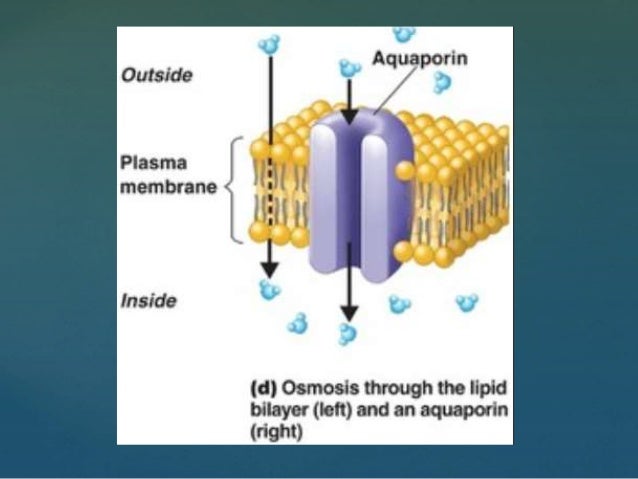
Несколько интересных и важных особенностей опосредованного аквапорином транспорта воды проиллюстрированы на основных клетках, которые выстилают собирательные трубочки в почках. Вода, текущая по этим протокам, может либо продолжаться и выводиться с мочой, либо реабсорбироваться через эпителий и обратно в кровь. Реабсорбция практически нулевая, если только эпителиальные клетки не реагируют на антидиуретический гормон, который сильно стимулирует реабсорбцию воды. Клетки собирательных протоков экспрессируют как минимум два аквапорина:
- Аквапорин-2 синтезируется, но в отсутствие антидиуретического гормона находится в пуле мембранных везикул в цитоплазме. Связывание антидиуретического гормона с его рецептором в клетке не только стимулирует транскрипцию гена аквапорина-2, но и вызывает встраивание внутриклеточного пула аквапорина-2 в апикальную мембрану. Теперь клетка способна эффективно поглощать воду из просвета протока.
- Аквапорин-3 конститутивно экспрессируется в базолатеральной мембране клетки.
 Когда вода попадает в клетку через каналы аквапорина-2, она может быстро выйти из клетки через каналы аквапорина-3 и попасть в кровь.
Когда вода попадает в клетку через каналы аквапорина-2, она может быстро выйти из клетки через каналы аквапорина-3 и попасть в кровь.
 Когда вода попадает в клетку через каналы аквапорина-2, она может быстро выйти из клетки через каналы аквапорина-3 и попасть в кровь.
Когда вода попадает в клетку через каналы аквапорина-2, она может быстро выйти из клетки через каналы аквапорина-3 и попасть в кровь.Аквапорины и заболевания
Принимая во внимание важность переноса воды во множестве физиологических процессов, следует ожидать, что повреждения генов аквапоринов или приобретенная дисфункция аквапоринов могут вызывать или способствовать возникновению нескольких болезненных состояний. Искать такие связи еще рано, но были идентифицированы два явных примера заболеваний, возникающих в результате дефицита аквапоринов:
- Мутации в гене аквапорина-2 вызывают у людей наследственный нефрогенный несахарный диабет.
- У мышей, гомозиготных по инактивирующей мутации в гене аквапорина-0, развивается врожденная катаракта.
- Ожирение у взрослых развивается у мышей, гомозиготных по инактивирующей мутации в гене аквапорина-7, предположительно из-за неэффективности транспорта глицерина из запасов гидролизованных триглицеридов.
У небольшого числа людей был выявлен тяжелый или полный дефицит аквапорина-1. Интересно, что они выглядят клинически незатронутыми, но не исследовались в условиях водной депривации. Мыши с целевыми делециями в аквапорине-1 также кажутся нормальными и здоровыми, если они не ограничены в жидкости, и в этом случае они становятся сильно гиперосмолярными.
Ссылки и обзоры
- Боргния М., Нильсен С., Энгель А., Агре П.: Клеточная и молекулярная биология аквапориновых водных каналов. Annu Rev Biochem 68:425, 1999.
- deGroot BL, Grubmuller H: Проникновение воды через биологические мембраны: механизм и динамика аквапорина-1 и GlpF. Наука 294:2353, 2001.
- Hibuse T, Proc Nat Acad Sci (США) 102:10993, 2005.
- Кинг Л.С., Агре П.: Патофизиология водных каналов аквапоринов. Анну Рев Физиол 58: 619, 1998.
- Кнеппер М.А., Иноуэ Т.: Регуляция водного канала аквапорина-2 с помощью вазопрессина. Curr Opinion Cell Biol 9:560, 1997.
- Lee MD, King LS, Agre P: Семейство аквапоринов белков водных каналов в клинической медицине. Медицина 76:141, 1997.
- Preston GM, Carroll TP, Guggino WP Agre P: Появление водных каналов в ооцитах Xenopus, экспрессирующих белок CHIP28 эритроцитов. Наука 256:385, 1992.
- Sasaki S, Ishibashi K, Marumo F: Аквапорины-2 и -3: представители двух подгрупп семейства аквапоринов, колокализованные в собирательных трубочках почек. Анну Рев Физиол 60:199, 1998.
Отправить комментарии по адресу [email protected]
Аквапорины – PMC
- Список журналов
- Рукописи авторов HHS
- PMC3590904
Карр Биол. Авторская рукопись; доступно в PMC 2014 21 января.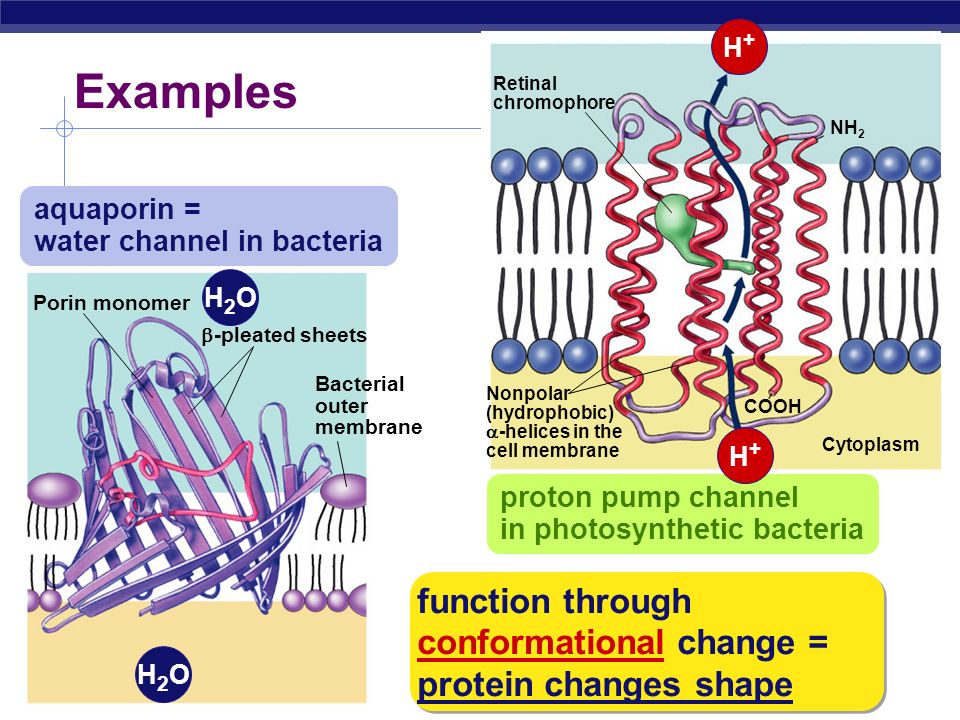
Опубликовано в окончательной редакции как:
Curr Biol. 2013 21 января; 23(2): Р52–Р55.
DOI: 10.1016/j.cub.2012.11.025
PMCID: PMC3590904
NIHMSID: NIHMS448095
PMID: 23347934
Информация о WATER COPYRIN небольших интегральных мембранных белков, которые широко экспрессируются во всех царствах животных и растений. Они имеют аналогичную базовую структуру с мономерами аквапоринов, состоящими из шести трансмембранных спиральных сегментов и двух коротких спиральных сегментов, которые окружают цитоплазматические и внеклеточные вестибюли, соединенные узкой водной порой. Они содержат несколько консервативных мотивов, включая последовательности NPA в своих коротких спиральных сегментах. Мономеры аквапорина собираются в виде тетрамеров в мембранах, причем каждый мономер функционирует независимо. Некоторые аквапорины, такие как AQP4 млекопитающих, могут дополнительно агрегировать в клеточных мембранах с образованием надмолекулярных кристаллических ансамблей, называемых ортогональными массивами частиц.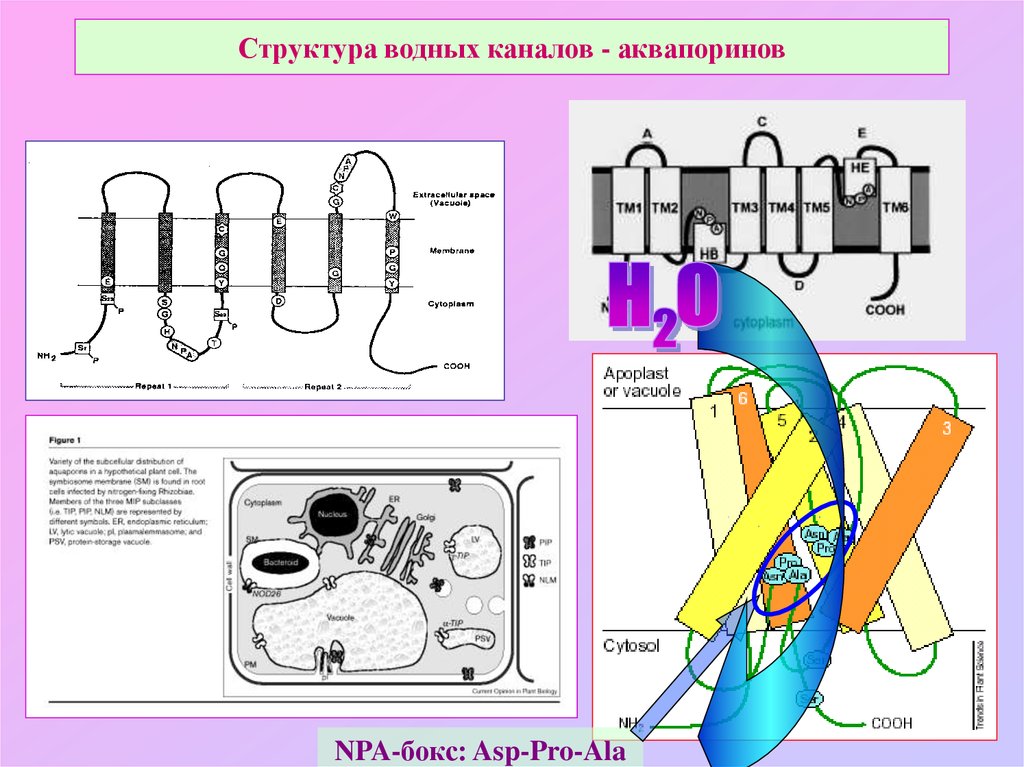
Открыть в отдельном окне
Структура и клеточные функции аквапоринов
(A) Мембранная топография мономера аквапорина (слева) и кристаллическая структура (AQP1, PDB 1j4n) (вид сбоку слева; вид сверху справа ) с четырьмя молекулами воды (красные шарики), показанными в водной области пор. Спирали помечены как h2–H8. (B) Секреция эпителиальной жидкости. Высокая трансэпителиальная водопроницаемость способствует активной почти изоосмолярной секреции жидкости экзокринными железами. (C) Миграция клеток. Миграция клеток, облегчаемая AQP, включает проникновение воды в выступающие ламеллиподии в мигрирующих клетках. (D) Увлажнение кожи. AQP3 поддерживает высокий уровень глицерина в роговом слое; глицерин действует как увлажнитель, удерживая воду. (E) Метаболизм адипоцитов. AQP7 способствует выходу глицерина из адипоцитов, предотвращая накопление внутриклеточного глицерина и триглицеридов. AQP, аквапорин; ТГ, триглицерид; СЖК, свободная жирная кислота.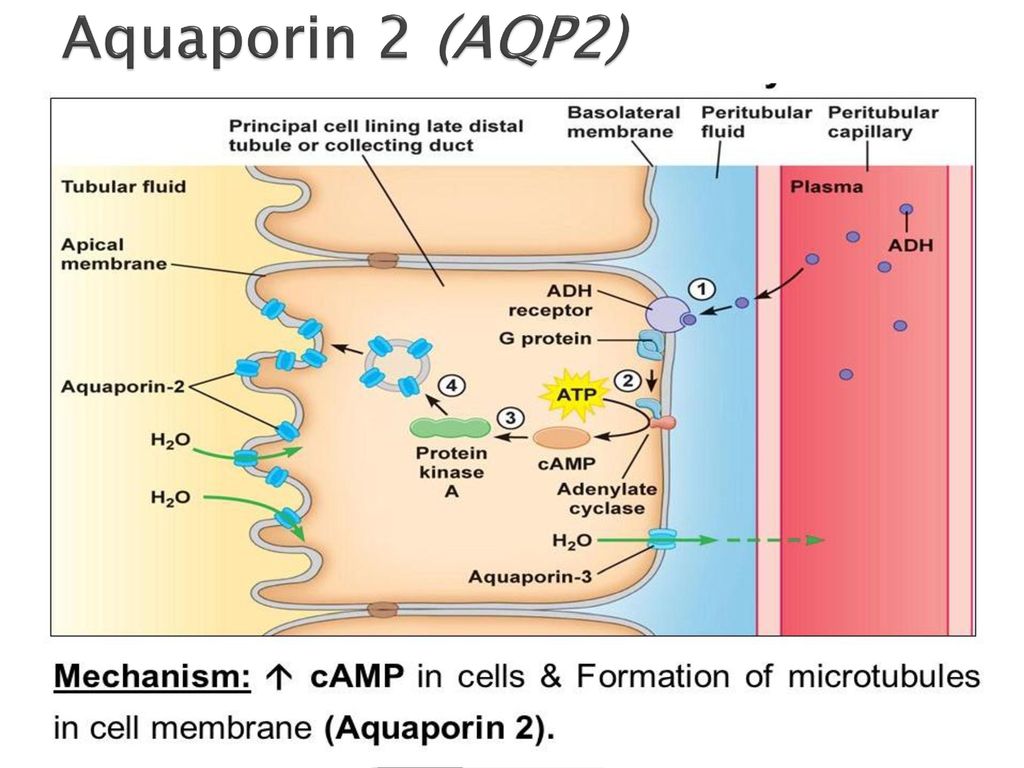 (Панели B–E адаптированы из Verkman, A.S. (2011). Аквапорины с первого взгляда. J. Cell Sci. 124 , 2107–2112.)
(Панели B–E адаптированы из Verkman, A.S. (2011). Аквапорины с первого взгляда. J. Cell Sci. 124 , 2107–2112.)
Основной функцией большинства аквапоринов является транспорт воды через клеточные мембраны в ответ на осмотические градиенты, создаваемые активным транспортом растворенных веществ. Поскольку водотранспортная способность мономеров аквапоринов низка, мембраны часто содержат аквапорины с высокой плотностью, до 10 000 на квадратный микрон, что значительно повышает водопроницаемость по сравнению с отсутствием аквапоринов. Моделирование молекулярной динамики предполагает, что стерические факторы и электростатические взаимодействия в водной поре ответственны за селективность аквапоринов по отношению к воде. Подгруппа аквапоринов, называемая акваглицеропоринами, также переносит глицерин. Диаметр пор акваглицеропоринов немного больше, чем у водоселективных аквапоринов, и поры выстланы относительно гидрофобными остатками по сравнению с порами водоселективных аквапоринов. Помимо воды и глицерина, есть доказательства, некоторые из которых противоречивы, что некоторые аквапорины пропускают газы (CO 2 , NH 3 , NO, O 2 ), различные мелкие растворенные вещества, такие как H 2 O 2 и арсенит, и даже ионы (K + , Cl – ). Также были предложены нетранспортные функции некоторых аквапоринов, такие как межклеточная адгезия, поляризация мембран и регуляция взаимодействующих белков, таких как ионные каналы.
Помимо воды и глицерина, есть доказательства, некоторые из которых противоречивы, что некоторые аквапорины пропускают газы (CO 2 , NH 3 , NO, O 2 ), различные мелкие растворенные вещества, такие как H 2 O 2 и арсенит, и даже ионы (K + , Cl – ). Также были предложены нетранспортные функции некоторых аквапоринов, такие как межклеточная адгезия, поляризация мембран и регуляция взаимодействующих белков, таких как ионные каналы.
Большинство клеток не экспрессируют аквапорины. Практически все биологические мембраны в достаточной степени проницаемы для воды вследствие диффузии воды через мембранные липиды, так что объем клетки уравновешивается за несколько минут или меньше в ответ на осмотический градиент. Так зачем же тогда в некоторых клетках нужна высокая водопроницаемость мембраны? Одним из примеров является секреция и абсорбция жидкости через слои эпителиальных клеток, например, в почечных канальцах и экзокринных железах, где нормальная функция почек и секреция телесных жидкостей, таких как слюна, требуют высокой трансэпителиальной проницаемости для воды, облегчаемой аквапорином. Высокая водопроницаемость также облегчает движение воды через барьеры между жидкостью и тканями, например, между кровью и паренхимой головного мозга через гематоэнцефалический барьер. Другие клеточные и тканевые функции водного транспорта аквапоринов и транспорта глицерина, облегчаемого акваглицеропорином, не очевидны априори .
Высокая водопроницаемость также облегчает движение воды через барьеры между жидкостью и тканями, например, между кровью и паренхимой головного мозга через гематоэнцефалический барьер. Другие клеточные и тканевые функции водного транспорта аквапоринов и транспорта глицерина, облегчаемого акваглицеропорином, не очевидны априори .
Аквапорины млекопитающих, которых насчитывается около дюжины, экспрессируются во многих типах клеток, участвующих в транспорте жидкости, включая эпителий и эндотелий в почках, легких, экзокринных железах, глазах и органах желудочно-кишечного тракта. Однако аквапорины также экспрессируются в клетках, не играющих очевидной роли в транспорте жидкости, таких как эритроциты и некоторые лейкоциты, адипоциты и скелетные мышцы. Аквапорины также экспрессируются в астроцитах по всей центральной нервной системе и в поддерживающих клетках (но не в электрически возбудимых клетках) в органах чувств, таких как клетки Мюллера сетчатки. В глазу аквапорины экспрессируются в роговице, хрусталике и цилиарном эпителии.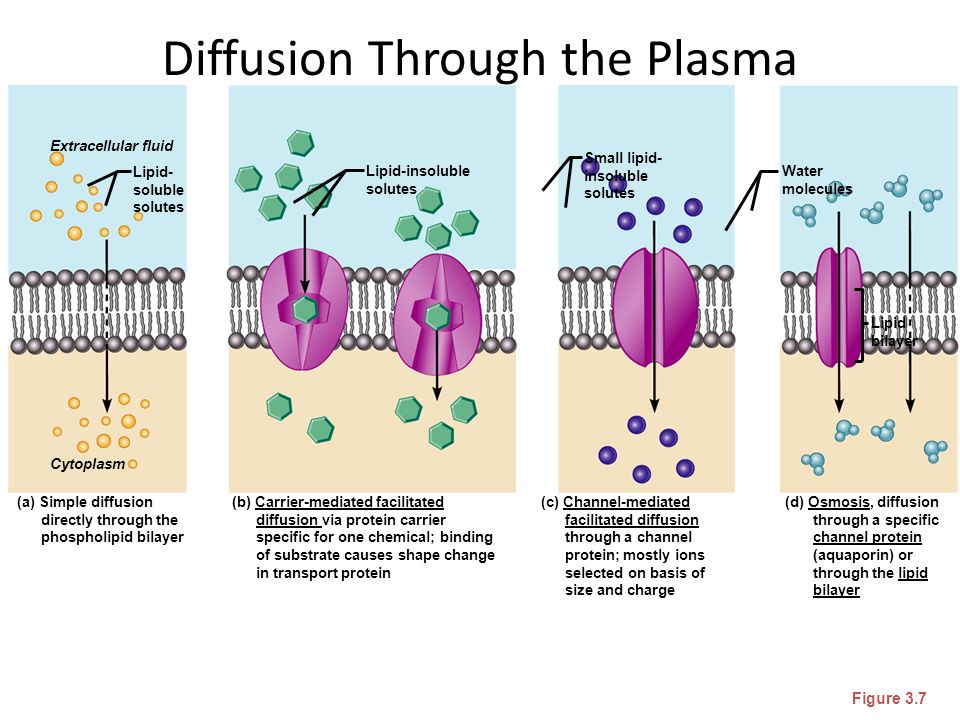 Распределение аквапоринов в тканях млекопитающих дало первоначальные сведения об их клеточных и органных функциях, хотя многие из первоначальных догадок не были подтверждены в исследованиях фенотипов на мышах с нокаутом по аквапоринам.
Распределение аквапоринов в тканях млекопитающих дало первоначальные сведения об их клеточных и органных функциях, хотя многие из первоначальных догадок не были подтверждены в исследованиях фенотипов на мышах с нокаутом по аквапоринам.
Аквапорины экспрессируются в различных растениях и микробах, включая бактерии и дрожжи. Было идентифицировано большое количество аквапоринов растений, названных PIP (внутренние белки плазматической мембраны) и TIP (внутренние белки тонопласта) из-за особенностей их клеточной экспрессии. Исследования делеций выявили множество интересных ролей аквапоринов в растениях, включая транспирацию, метаболизм и репродукцию, которые важны для адаптации растений к различным стрессам окружающей среды. Хотя аквапорины также экспрессируются в некоторых микробах, исследования делеций не выявили какого-либо четкого фенотипа, хотя есть наводящие доказательства того, что аквапорины могут быть защитными при стрессе замораживания-оттаивания, возможно, для поддержания водопроницаемости клеточных мембран при низкой температуре. Высокое отношение поверхности к объему микробов предсказывает быстрое осмотическое уравновешивание, что делает аквапорины ненужными для большинства функций, таких как осморегуляция.
Высокое отношение поверхности к объему микробов предсказывает быстрое осмотическое уравновешивание, что делает аквапорины ненужными для большинства функций, таких как осморегуляция.
Фенотипический анализ мышей с нокаутом по аквапоринам подтвердил ожидаемую роль аквапоринов в трансэпителиальном транспорте жидкости (). Удаление различных аквапоринов в почечных канальцах вызывает избыточный диурез и нарушение концентрационной способности мочи. Высокая водопроницаемость, зависящая от аквапоринов, необходима в проксимальных канальцах почек для почти изоосмолярной абсорбции жидкости, в тонком нисходящем колене Генле для противотока и в собирательных трубочках для абсорбции воды. Аквапорины также способствуют секреции эпителиальной жидкости в слюнных и подслизистых железах дыхательных путей, а также в сосудистом сплетении (из которого секретируется спинномозговая жидкость) и цилиарном эпителии (из которого секретируется глазная жидкость). Однако есть важное предостережение: аквапорины необходимы для трансэпителиального транспорта жидкости, когда скорость транспорта жидкости (на площадь эпителиальной поверхности) очень высока, как в проксимальных канальцах и слюнных железах. Хотя аквапорины экспрессируются и функционируют в эпителии легких, кишечника и потовых желез, они, по-видимому, не играют значительной физиологической роли.
Хотя аквапорины экспрессируются и функционируют в эпителии легких, кишечника и потовых желез, они, по-видимому, не играют значительной физиологической роли.
Непредвиденная роль аквапоринов заключается в миграции клеток. Делеция AQP1 у мышей нарушает ангиогенез как следствие снижения скорости миграции эндотелиальных клеток микрососудов. Аквапорин-зависимая миграция была обнаружена в самых разных типах клеток. Биофизические исследования предполагают, что поляризация аквапоринов к переднему краю мигрирующих клеток облегчает приток воды во время ламеллоподиального растяжения (4). В отличие от трансэпителиального транспорта жидкости, где аквапорины облегчают движение воды через клеточные слои, аквапорины в мигрирующих клетках облегчают локальный транзиторный транспорт воды. Другая непредвиденная роль аквапорина заключается в нейровозбуждении: делеция AQP4 ухудшает нейросенсорную передачу сигналов и изменяет динамику судорог и распространяющейся в коре депрессии. Хотя этот механизм до конца не выяснен, похоже, что транспорт воды, облегчаемый AQP4, необходим для быстрых изменений объема внеклеточного пространства и K + , и что потеря AQP4 приводит к замедленному обратному захвату K + после нейровозбуждения.![]() Другие неожиданные роли водоселективных аквапоринов включают их участие в прозрачности роговицы и хрусталика, отек головного и спинного мозга, нейровоспаление и боль.
Другие неожиданные роли водоселективных аквапоринов включают их участие в прозрачности роговицы и хрусталика, отек головного и спинного мозга, нейровоспаление и боль.
Акваглицеропорины млекопитающих включают AQP3, AQP7 и AQP9. AQP3 экспрессируется в базальном слое пролиферирующих кератиноцитов эпидермиса. У мышей, лишенных AQP3, снижена гидратация и эластичность кожи, что связано с нарушением транспорта глицерина из крови в эпидермис и, как следствие, снижением содержания глицерина в эпидермисе и роговом слое. Уменьшение содержания глицерина ухудшает увлажнение и эластичность кожи, так как глицерин является основным увлажнителем (осмолитом, удерживающим воду). Снижение уровня глицерина также нарушает эпидермальную пролиферацию, поскольку глицерин является важным метаболитом эпидермальных клеток, участвующим в образовании АТФ и биосинтезе мембранных липидов. В косметической промышленности наблюдается значительный интерес к AQP3, при этом заявленные на рынке продукты повышают экспрессию AQP3.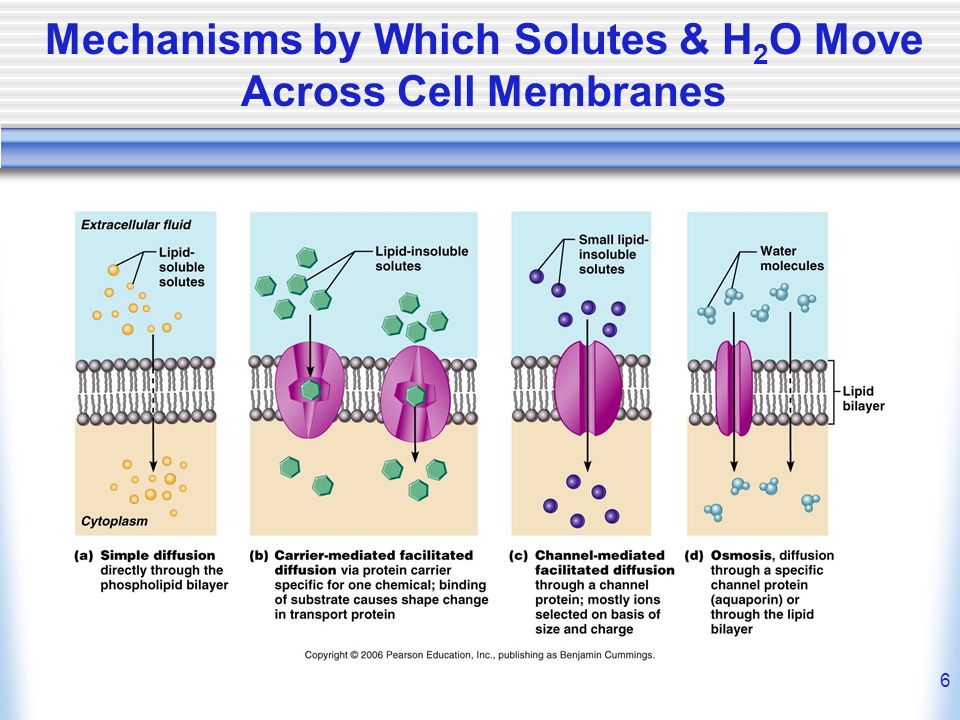 Недавние исследования показали важную роль AQP3 в функции макрофагов и Т-лимфоцитов, возможно, с участием AQP3-облегченного H9.0269 2 O 2 транспорт. Другое интригующее наблюдение, сделанное на мышах с недостатком AQP7, представляет собой прогрессирующее ожирение с гипертрофией адипоцитов, которое, по-видимому, является следствием нарушения оттока глицерола из адипоцитов. Снижение проницаемости плазматической мембраны для глицерина в результате дефицита AQP7 приводит к внутриклеточному накоплению глицерина и триглицеридов (2), что указывает на возможность активации AQP7 для уменьшения жировой массы при ожирении. Наконец, предполагается, что экспрессия AQP9 в гепатоцитах способствует поглощению глицерина печенью, хотя значение поглощения глицерина печенью в метаболизме человека неясно.
Недавние исследования показали важную роль AQP3 в функции макрофагов и Т-лимфоцитов, возможно, с участием AQP3-облегченного H9.0269 2 O 2 транспорт. Другое интригующее наблюдение, сделанное на мышах с недостатком AQP7, представляет собой прогрессирующее ожирение с гипертрофией адипоцитов, которое, по-видимому, является следствием нарушения оттока глицерола из адипоцитов. Снижение проницаемости плазматической мембраны для глицерина в результате дефицита AQP7 приводит к внутриклеточному накоплению глицерина и триглицеридов (2), что указывает на возможность активации AQP7 для уменьшения жировой массы при ожирении. Наконец, предполагается, что экспрессия AQP9 в гепатоцитах способствует поглощению глицерина печенью, хотя значение поглощения глицерина печенью в метаболизме человека неясно.
Провокационные данные указывают на причастность аквапоринов к некоторым видам рака. В большом количестве описательной литературы сообщается о сильной экспрессии аквапоринов при раке головного мозга, кожи, желудочно-кишечного тракта, легких и других видах рака, часто с корреляцией между экспрессией аквапоринов и степенью опухоли. Опухоли часто сверхэкспрессируют аквапорины, обнаруженные в типе клеток происхождения, таких как AQP4 в глиобластомах, происходящих из астроцитов, хотя иногда опухолевые клетки экспрессируют аквапорины, которые не наблюдаются в клетках происхождения. Было предложено несколько механизмов, которые связывают экспрессию аквапоринов с раком. AQP1 сильно экспрессируется в микрососудах опухоли, и его делеция у мышей снижает ангиогенез опухоли. Экспрессия различных аквапоринов в опухолевых клетках увеличивает их миграцию и метастатический потенциал в клеточных культурах и животных моделях. AQP3 увеличивает пролиферацию различных опухолевых клеток, а мыши, лишенные AQP3, обладают высокой устойчивостью к раку кожи. Хотя многие вопросы еще предстоит решить, ингибирование аквапорина для лечения рака является интригующей идеей.
Опухоли часто сверхэкспрессируют аквапорины, обнаруженные в типе клеток происхождения, таких как AQP4 в глиобластомах, происходящих из астроцитов, хотя иногда опухолевые клетки экспрессируют аквапорины, которые не наблюдаются в клетках происхождения. Было предложено несколько механизмов, которые связывают экспрессию аквапоринов с раком. AQP1 сильно экспрессируется в микрососудах опухоли, и его делеция у мышей снижает ангиогенез опухоли. Экспрессия различных аквапоринов в опухолевых клетках увеличивает их миграцию и метастатический потенциал в клеточных культурах и животных моделях. AQP3 увеличивает пролиферацию различных опухолевых клеток, а мыши, лишенные AQP3, обладают высокой устойчивостью к раку кожи. Хотя многие вопросы еще предстоит решить, ингибирование аквапорина для лечения рака является интригующей идеей.
Одно редкое наследственное заболевание вызывается мутациями потери функции в AQP2, водном канале, экспрессирующемся в собирательных трубочках почек. AQP2 способствует трансэпителиальному всасыванию воды в собирательных трубочках почек в ответ на антидиуретический гормон, и этот процесс включает экзоцитарную вставку AQP2-содержащих везикул в апикальную мембрану клетки. Мутации в AQP2, которые нарушают его клеточный транспорт или функцию транспорта воды, приводят к заболеванию нефрогенным несахарным диабетом, при котором выделяются большие объемы разбавленной мочи, иногда превышающие 30 литров в день. Мутации в AQP0 (первоначально названном «основным внутренним белком волокон хрусталика») связаны с врожденными катарактами; однако механизм катарактогенеза неизвестен, поскольку неясно, заключается ли основная функция AQP0 в структуре хрусталика или в водопроницаемости клеток волокон хрусталика. Мутации потери функции в нескольких других аквапоринах были идентифицированы у небольшого числа людей, хотя ни одна из них, по-видимому, не связана со значительным заболеванием. Интересное аутоиммунное неврологическое заболевание, оптиконейромиелит, вызывается аутоантителами, направленными против внеклеточных эпитопов на AQP4. Связывание аутоантител вызывает комплемент- и клеточно-опосредованную цитотоксичность астроцитов, что приводит к воспалению с высвобождением цитокинов, инфильтрацией лейкоцитов и нарушением гематоэнцефалического барьера.
Мутации в AQP2, которые нарушают его клеточный транспорт или функцию транспорта воды, приводят к заболеванию нефрогенным несахарным диабетом, при котором выделяются большие объемы разбавленной мочи, иногда превышающие 30 литров в день. Мутации в AQP0 (первоначально названном «основным внутренним белком волокон хрусталика») связаны с врожденными катарактами; однако механизм катарактогенеза неизвестен, поскольку неясно, заключается ли основная функция AQP0 в структуре хрусталика или в водопроницаемости клеток волокон хрусталика. Мутации потери функции в нескольких других аквапоринах были идентифицированы у небольшого числа людей, хотя ни одна из них, по-видимому, не связана со значительным заболеванием. Интересное аутоиммунное неврологическое заболевание, оптиконейромиелит, вызывается аутоантителами, направленными против внеклеточных эпитопов на AQP4. Связывание аутоантител вызывает комплемент- и клеточно-опосредованную цитотоксичность астроцитов, что приводит к воспалению с высвобождением цитокинов, инфильтрацией лейкоцитов и нарушением гематоэнцефалического барьера.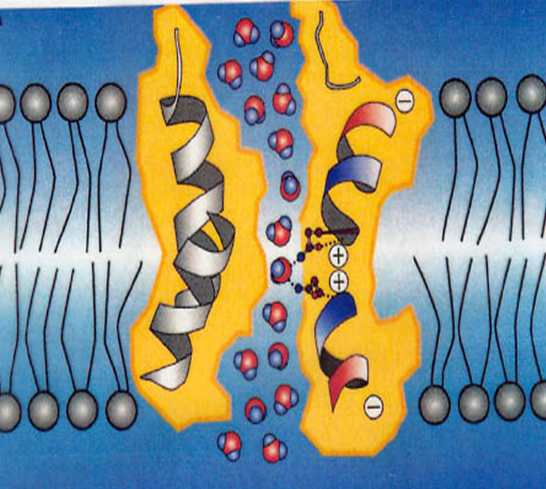 Воспалительные демиелинизирующие поражения спинного мозга и зрительного нерва приводят к параличу и слепоте. До сих пор не были обнаружены другие аутоиммунные заболевания, основанные на аквапорине.
Воспалительные демиелинизирующие поражения спинного мозга и зрительного нерва приводят к параличу и слепоте. До сих пор не были обнаружены другие аутоиммунные заболевания, основанные на аквапорине.
Существует значительный интерес, хотя и небольшой прогресс в области терапии на основе аквапоринов. Данные клеточных и животных моделей обеспечивают убедительное обоснование разработки модуляторов аквапоринов в качестве терапевтических средств: предполагается, что ингибиторы AQP1 будут полезны при рефрактерных к диуретикам отеках, опухолевом ангиогенезе и глаукоме; Ингибиторы AQP4 при некоторых формах отека и травм головного мозга; и ингибиторы AQP3 при некоторых видах рака. Повышающие регуляторы экспрессии некоторых аквапоринов имеют потенциальное терапевтическое значение при ожирении, заживлении ран и повреждении спинного мозга. Определенные соединения тяжелых металлов (ртуть, золото), которые неспецифически реагируют с сульфгидрилами белков, ингибируют водопроницаемость некоторых аквапоринов.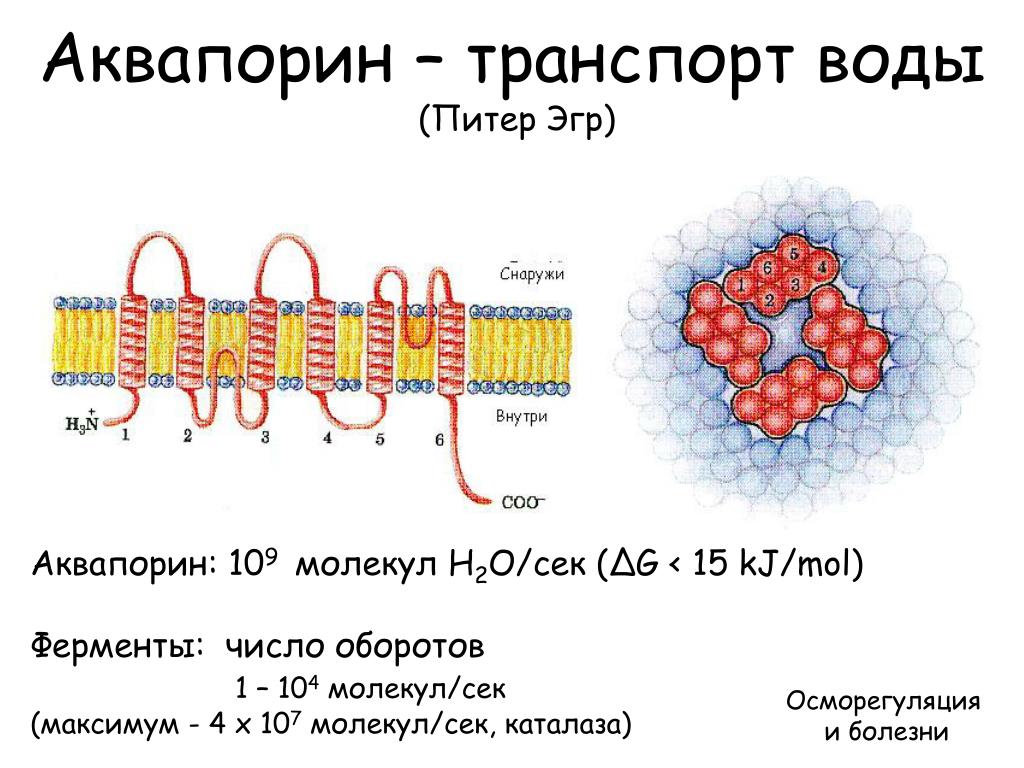 Тем не менее, идентификация нетоксичных низкомолекулярных ингибиторов аквапоринов оказалась невозможной, возможно, по совокупности причин, включая трудности в функциональных скрининговых анализах и присущую аквапоринам невосприимчивость к открытию лекарств (так называемая нелекарственность). Одной из областей недавнего прогресса является терапия на основе AQP4 для лечения оптикомиелита. Идея состоит в том, чтобы заблокировать связывание патогенных аутоантител с астроцитом AQP4, что может обеспечить целенаправленный неиммуносупрессивный подход к блокированию события, инициирующего заболевание, при оптикомиелите. Одним из подходов была разработка непатогенного, прочно связывающегося антитела против AQP4 («аквапорумаб»), которое конкурирует с патогенными аутоантителами; другим подходом была идентификация с помощью высокопроизводительного скрининга низкомолекулярных блокаторов взаимодействия аутоантитело-AQP4.
Тем не менее, идентификация нетоксичных низкомолекулярных ингибиторов аквапоринов оказалась невозможной, возможно, по совокупности причин, включая трудности в функциональных скрининговых анализах и присущую аквапоринам невосприимчивость к открытию лекарств (так называемая нелекарственность). Одной из областей недавнего прогресса является терапия на основе AQP4 для лечения оптикомиелита. Идея состоит в том, чтобы заблокировать связывание патогенных аутоантител с астроцитом AQP4, что может обеспечить целенаправленный неиммуносупрессивный подход к блокированию события, инициирующего заболевание, при оптикомиелите. Одним из подходов была разработка непатогенного, прочно связывающегося антитела против AQP4 («аквапорумаб»), которое конкурирует с патогенными аутоантителами; другим подходом была идентификация с помощью высокопроизводительного скрининга низкомолекулярных блокаторов взаимодействия аутоантитело-AQP4.
В наших знаниях о клеточных механизмах некоторых функций аквапоринов, таких как миграция клеток, облегчаемая аквапорином, пролиферация клеток и явления нейровозбуждения, остаются значительные пробелы.
Добавить комментарий